На главную
ЧАСТЬ ПЕРВАЯ
Известно, что:
-яичник в репродуктивной системе является филогенетически самым древним органом;
-эстрогены обнаруживаются практически у всех представителей животного мира;
-фолликулярный оогенез характерен для многих безпозвоночных, всех позвоночных и человека;
-остальные органы репродуктивной системы являются мишенями, реагирующими на стероидные гормоны.
Возникает ряд вопросов:
-являются ли стероидные гормоны веществами, специально выделяющимися в яичнике для влияния на органы-мишени, более молодые в филогенетическом отношении?
-или же органы мишени реагируют на продукты жизнедеятельности яичника (по Barrington ' y - адаптация тканевых структур к предшествующему гормону)?
-если это так, то какие же продукты и каких процессов являются специфическими только для яичников?
Известно, что в яичнике происходит:
-оогенез, заключающийся в росте и созревании ооцита до стадии, способной к оплодотворению и дальнейшему развитию;
-фолликулогенез, заключающийся на первых этапах в активной пролиферации гранулезы.
После овуляции и до имплантации ооцит находится практически в автономном существовании и в это время с ним происходит ряд процессов (оплодотворение, дробление и формирование бластоцисты).
Известно также, что:
-бластоциста в предимплантационной стадии состоит из сотен клеток;
-в состав клеточных оболочек, которые являются биологическими мембранами, входят липиды, в том числе и соединения холестерина;
-биосинтез эстрогенов в яичнике представляет из себя линейный процесс: холестерин - прогестерон - андрогены - эстрогены;
- вектор этого процесса направлен внутрь фолликула: тека-гранулеза;
-концентрация эстрадиола в фолликулярной жидкости до 1000 раз превышает концентрацию в периферической крови;
- на первых фазах развития фолликула диаметр ооцита увеличивается в 10 раз.
Учитывая вышеизложенное, можно сделать предположение, что для образования в период от оплодотворения до стадии бластоцисты большого количества биологических мембран необходимо большое количество белков и липидов, запас последних в ооците создается с помощью клеток теки и гранулезы, в которых холестерин превращается в более легко усвояемую форму — эстрадиол.
Изучая физиологию системы ооцит - фолликул можно сделать вывод о том, что существуют два механизма: механизм созревания ооцита и механизм изгнания ооцита. В норме эти два процесса происходят синхронно.
Известно, что:
-между текой и гранулезой находится базальная мембрана;
-все клетки гранулезы соприкасаются отростками с базальной мембраной и ооцитом;
-скорость развития фолликула до стадии полости довольно постоянная и не зависит от гормонов гипофиза;
- рост ооцита происходит без влияния гонадотропных гомонов;
-пока ооцит не закончит рост, в фолликулярных клетках не происходит существенных изменений и фолликул остается компактным;
-после роста ооцита в фолликулярных клетках наблюдается выраженная митотическая активность;
-появление полости в фолликуле совпадает по времени с началом развития внутренней теки;
- характерная особенность внутренней теки — богатая васкуляризация;
- фолликулярная жидкость человека обладает резко выраженным митогенным эффектом в отношении эндотелиальных клеток. Наблюдается направленная миграция(хемотаксис) эндотелиальных клеток к фолликулярной жидкости человека. Она об ладает ангиогенным действием;
- в фолликуле существует гисто-гематический (или гемато-фолликулярный) барьер, основная структурная единица которого — базальная мембрана и функции его - высокое сопротивление диффузному потоку и избирательная проницаемость.
Учитывая вышеизложенное, можно рассуждать следующим образом. В растущем фолликуле наряду с ростом ооцита происходит пролиферация гранулезы. Как пролиферирующая ткань гранулеза вырабатывает вещества, стимулирующие неоваскулогенез. Но базальная мембрана не пропускает вновь образованные сосуды внутрь фолликула, поэтому они концентрируются по периферии фолликула на поверхности мембраны, образуя вместе с привнесенными интерстициальными клетками внутреннюю теку. С появлением сосудов и образованием теки ФСГ начинает действовать на гранулезу. Развития гранулезы до образования полости достаточно для созревания ооцита, о чем свидетельствует остановка роста ооцита. В этот момент включается механизм изгнания ооцита. Под действием ФСГ гранулеза продолжает пролиферировать. Образование полости - естестественное следствие гиперпролиферации гранулезы. Некоторые клетки гранулезы теряют контакт с ооцитом. Продолжающаяся секреция этими клетками приводит к накапливанию внутри фолликула большого количества эстрадиола.
Известно, что:
- эстрадиол ингибирует секрецию гонадотропинов, подавляя секрецию гонадолиберина гипоталамусом;
- секреция как гонадолиберина, так и гонадотропинов носит импульсный характер с интервалом около часа;
- аденогипофиз не имеет прямых нейротропных связей с гипоталамусом;
- период полураспада ЛГ - 12-45 мин., ФСГ - 12 мин;
- запасы ЛГ и ФСГ в гипофизе во много раз превышают их концентрацию в периферической крови - регистрируемые изменения уровня ЛГ в крови практически не отражают содержания ЛГ в гипофизе;
-введение прогестерона урежает частоту импульсов гонадотропинов, но увеличивает выброс, а введение эстрадиола уменьшает количество выброса их до десяти раз.
Учитывая вышеизложенное, можно сделать вывод, что секреция гонадотропинов носит импульсный характер, вероятно, в связи с изменениями концентрации легкораспадающихся гонадотропинов в крови. Концентрация гонадотропинов в крови, достигая определенной величины, блокирует выделение гонадолиберина гипоталамусом. Секреция гонадотропинов прекращается. Но в связи с коротким периодом жизни концентрация гонадотропинов в крови снижается, блок в гипоталамусе снимается и выделяющийся гонадолиберин вновь стимулирует высвобождение гонадотропинов из гонадотрофов. Таким образом осуществляется ауторегуляция импульсной секреции гонадотропных гармонов. Доказательством этому может быть тот факт, что хорионический гонадотропин, являясь аналогом ЛГ (выделение ХГ при беременности не зависит от гипоталамуса и период полураспада его 24 часа), блокирует стимуляцию выделения гонадотропинов, тем самым предотвращает рост и созревание фолликулов. Для воздействия на яичники не обязателен импульсный характер выделения гонадотропных гормонов. Доказательством этому служит пример эффективности применения препаратов менопаузального гонадотропина.
Половые гормоны оказывают модулирующее влияние на характер пульсаций выбросов.
Известно, что:
- ооцит в примордиальном фолликуле находится в диктиотене профазы мейоза;
- ко времени овуляции в ооците возобновляется процесс мейоза: заканчивается первое деление созревания. Отделяется полярное тельце и ооцит переходит во второе деление созревания;
- длительность предовуляционного нарастания уровня эстрадиола в периферической крови колеблется в узких пределах - 4-6 дней, причем продолжительность этого периода не зависит от общей продолжительности фолликулиновой фазы цикла;
- в созревающем фолликуле гранулезный эпителий подвергается митотическому распаду, в результате чего образуется фолликулярная жидкость. Гранулезная оболочка предовуляторного фолликула у человека состоит из 2-3 рядов клеток;
- стенка фолликула перед овуляцией содержит гладкомышечные клетки.
Исходя из вышеперечисленного можно предположить, что ооцит в процессе созревания проходит такую стадию или фазу, во время которой выделяются вещества, лизирующие основную массу гранулезы. Остаются лишь клетки, тесно контактирующие или с ооцитом или с базальной мембраной. В фолликулярную жидкость выбрасывается большое количество эстрадиола — мерокриновая секреция сменяется голокриновой. Кроме эстрадиола, освобождается большое количество ферментов, в том числе протеиназы, гиалуронидазы. Разрушается базальная мембрана. Капилляры начинают прорастать внутрь фолликула, оплетая каждую клетку гранулезы. Препятствие диффузному потоку в виде гемато-фолликулярного барьера устраняется. Концентрация эстрадиола в периферической крови резко увеличивается (т. к. концентрация его в фолликуле превышала концентрацию в периферической крови до 1000 раз).
Высокая концентрация эстрадиола блокирует выработку гонадолиберина гипоталамусом. Но период полураспада гормонов очень короткий ( 5-20 мин). А гранулезные клетки больше не секретируют эстрадиол. Запас эстрадиола в фолликулярной жидкости иссякает, концентрация его в крови падает и блок в гипоталамусе снимается. Как реакция на отмену блока система гипоталамус - гипофиз отвечает выбросом гонадотропинов. В результате этого выброса запасы секрета в гонадотрофах истощаются и уровень гонадотропинов в крови в дальнейшем резко снижается. Доказательством существования эффекта «истощения» гонадотрофов служит пример применения синтетических аналогов гонадолиберина — золадекса и др.: после введения имплантанта постоянное выделение препарата вызывает вначале повышение содержания гонадотропинов в крови, а затем снижение их до постменопаузального уровня.
Падение уровня гонадотропинов снимает блок с гипоталамуса и стимулирует в свою очередь высвобождение окситоцина в задней доле гипофиза.
Высокий же уровень гонадотропинов (ЛГ) во время пика, воздействуя на оставшиеся клетки гранулезы, превращает их в настоящие стероидпродуцирующие клетки.
В фолликуле в связи с высокой концентрацией веществ и разрушением барьера в полость стремительно поступает жидкость. Фолликул за несколько часов резко увеличивается в объеме. Давление на стенки возрастает, о чем свидетельствует быстрый рост объема жидкости и сферическая форма фолликула. Кровообращение его стенки нарушается, о чем свидетельствуют кровоизлияния в стенку. Та часть стенки, которая ближе к поверхности яичника, истончается, капилляры истончаются и запустевают. Локальная гипоксия приводит к гибели клеток и освобождению протеолитических ферментов, оказывающих местное воздействие. Окситоцин, выделяющийся перед овуляцией, стимулируя синтез простагландинов, воздействует на гладкомышечные клетки стенки фолликула, повышает давление на содержимое фолликула и способствует овуляции. Ооцит выходит в брюшную полость с током жидкости вследствие сокращения стенок фолликула.
Рассогласование двух процессов - механизма созревания ооцита и механизма изгнания его - приводит к появлению таких нарушений, как атрезия фолликула, персистенция фолликула, «перезревание» ооцита, овуляция дегенеративным ооцитом, лютеинизация неразорвавшегося фолликула (встречающаяся у здоровых фертильных женщин, по разным авторам, от 9 до 55%). Этим же объясняется механизм действия оральных контрацептивов.
После овуляции на месте фолликула образуется желтое тело из клеток внутренней теки и оставшихся гранулезных клеток, которые находятся как бы в парабиозе — отсутствуют митозы, синтез стероидов останавливается на прогестероне, ароматазная активность выключается, секрет накапливается в клетках, увеличивая их в размерах. Выделяющийся в больших количествах прогестерон блокирует повышение уровня гонадотропных гормонов. Яичник, образно говоря, «замер в ожидании». Долго в таком состоянии клетки находится не могут и при отсутствии оплодотворения ооцита в клетках желтого тела начинается процесс дегенерации. Снижение уровня прогестерона в конце II фазы приводит к разблокировке гипоталамуса и, возможно, является концом гормоннезависимого периода развития примордиальных фолликулов и началом действия механизма изгнания следующего ооцита.
После оплодотворения ооцита развивающаяся бластоциста еще в предимплантационной стадии начинает продуцировать ХГ. Являясь аналогом ЛГ, он стимулирует пролиферацию гранулезолютеиновых клеток и секрецию прогестерона. Блок гипоталамуса осуществляется как высокой концентрацией половых гомонов, так и все увеличивающейся концентрацией ХГ. Концентрация ХГ в периферической крови растет до 11- 12 недели беременности. Затем после начала функционирования плаценты блок гипоталамуса осуществляется высоким содержанием гормонов, синтезируемых в фето-плацентарном комплексе (существует также и местный блок).
Вот почему аксоны нейронов гипоталамуса заканчиваются в задней доле гипофиза, т.е. при любом снятии блока гипоталамуса увеличивается выделение окситоцина: перед овуляцией, перед менструацией, при угрожающем аборте, перед родами.
Таким образом можно сделать вывод о том, что звено гипотамус - гипофиз предназначено для изгнания продуктов системы - яйцеклетки и плодного яйца. Гонадотропные гормоны, обеспечивая механизм изгнания яйцеклетки, являются сдерживающим фактором в этой системе, определяют в силу своей короткой жизни пульсирующий характер выделеления гонадолиберина. Пульсирующий характер необходим для обеспечения запаса гормонов в гонадотрофах. Половые гормоны как продукты жизнедеятельности фолликула и плодного яйца, осуществляют блокирующее воздействие на это звено, определяя в конце концов время активизации изгоняющей системы.
Во всей репродуктивной системе осуществляется принцип «блокады» гипоталамуса будь то фолликул, желтое тело, гонадотрофы или плодное яйцо.
ЧАСТЬ ВТОРАЯ
Иерархическая теория регуляции предполагает участие коры головного мозга в регуляции репродуктивной системы женщины. Так ли это? Существуют открытые и закрытые системы. Более надежные — закрытые, саморегулирующиеся. У низших животных репродуктивные системы открытые, т.е. предусматривается участие коры головного мозга: у них функционирование системы зависит от времени года, времени суток, температуры окружающей среды, контакта с половым партнером и др. У человека же как высшего существа репродуктивная система более надежна и не зависит от внешних условий, т.е. закрытая, в регуляции которой не подразумевается участие коры головного мозга. Влияние коры носит скорее возмущающий характер, чем регулирующий. Что же происходит в гипоталамо-гипофизарном звене?
Известно, что:
- секреция пролактина ( ПЛ ) блокируется дофамином;
- секреция гонадотропин-рилизинг-гормона (ГнРГ) блокируется опиоидными пептидами;
- секреция ПЛ стимулируется опиоидными пептидами посредством торможения дофаминергической системы;
- ГнРГ стимулирует секрецию лютеинизирующего гормона (ЛГ);
-дофамин не оказывает непосредственного эффекта на секрецию ЛГ, а стимулирует высвобождение опиоидных пептидов из гипоталамуса и тормозит секрецию ЛГ;
- опиоидные пептиды содержатся в нейронах вместе с нейрогипофизарными гормонами (окситоцином и вазопрессином) и тормозят их выделение;
- после падения опиоидной активности следует увеличение простагландиновой активности;
- торможение выделения ГнРГ опиоидными пептидами модулируется овариальными стероидами;
- гипоталамический ? - эндорфин секретируется в портальную систему, и эта секреция увеличивается на фоне лечения эстрогенами;
• в промежуточной доле гипофиза, но не в головном мозге, ? — эндорфин подвергается ? — N - ацетилированию, приводящему к полной потере опиатоподобной активности пептида.
Исходя из вышеизложенного, можно сделать вывод о том, что опиоидные пептиды являются в гипоталамусе универсальными ингибиторами: тормозят секрецию ГнРГ, дофамина и окситоцина и секреция этих пептидов стимулируется эстрогенами.
Выше было высказано предположение об ауторегуляции импульсной секреции ГнРГ и ЛГ на основании данных о коротком периоде полураспада ЛГ в крови.
На основании всего вышеизложенного гипоталамо-гипофизарные отношения можно предположить в следующем виде (см. рис. 1).
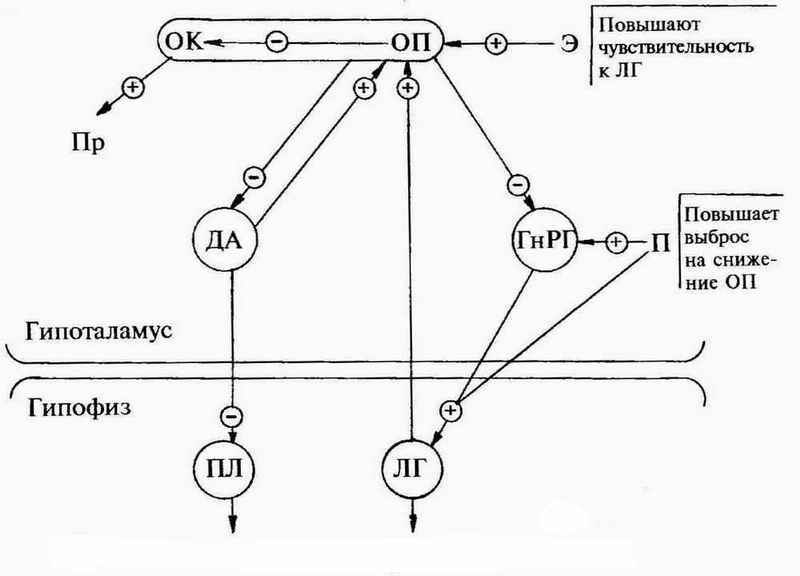
Рис 1: + -стимулирующее влияние, - -тормозящее влияние
Согласно этой схемы пульсирующий характер секреции ГнРГ и ЛГ можно объяснить следующим образом: секретирующиеся в паравентрикулярных и супраоптических ядрах гипоталамуса опиоидные пептиды (ОП) тормозят секрецию в медиобазальном гипоталамусе (в дугообразном ядре) ГнРГ. При снижении секреции ОП до определенного уровня начинается секреция ГнРГ, под действием которого в гонадотрофах гипофиза начинается секреция ЛГ. ЛГ оказывает стимулирующее воздействие на секрецию ОП гипоталамусом, попадая туда обратным током крови из портальной системы. Под действием ЛГ уровень ОП повышается и оказывает блокирующее воздействие на медиобазальный гипоталамус. Секреция ГнРГ прекращается. Соответственно прекращается выброс ЛГ. Но ЛГ легкораспадающийся гормон, через некоторое время концентрация его в крови снижается, секреция ОП снижается, снимается блок с медиобазального гипоталамуса и возобновляется секреция ГнРГ. Цикл повторяется.
Эстрогены оказывают стимулирующее воздействие на секрецию ОП путем повышения чувствительности к ЛГ. То есть для начала повышения секреции ОП становится достаточно меньшего уровня ЛГ, чем без эстрогенов. Таким образом снижается порог чувствительности к ЛГ и этот порог тем ниже, чем выше концентрация эстрогенов и в конце концов может привести к полному блоку секреции ГнРГ, что и происходит при лечении большими дозами эстрогенов. Этим феноменом объясняется также механизм действия эстрогеновых контрацептивов.
Стимулирующим воздействием на секрецию ОП при лечении эстрогенами объясняется гиперпролактинемия, что клинически проявляется в нагрубании молочных желез: повышение секреции ОП ведет к блокаде дофаминергической системы, что, в свою очередь, снимает блок с лактотрофов в гипофизе.
Прогестерон же, по всей видимости, оказывает стимулирующее воздействие на медиобазальный гипоталамус, повышает чувствительность секреции ГнРГ на снижение уровня ОП, что проявляется увеличением выброса ГнРГ и, как следствие, увеличением выброса ЛГ. Естественно повышенная концентрация ЛГ в крови оказывает более длительное стимулирующее воздействие на секрецию ОП, пока концентрация ЛГ не снизится до уровня, когда секреция ОП начнет снижаться. Этим объясняется тот факт, что во II фазу цикла амплитуда выбросов ЛГ увеличивается с одновременным снижением частоты.
В середине цикла в связи с предовуляторными изменениями в яичнике концентрация эстрогенов в крови резко повышается, что приводит к блоку секреции ГнРГ и ЛГ. Следующее за пиком резкое снижение концентрации эстрогенов приводит к снижению секреции опиоидных пептидов и разблокировке секреции ГнРГ, что проявляется освобождением из гонадотрофов гипофиза большого количества ЛГ.
Перед менструацией низкий уровень Э и резкое снижение П приводят к снижению секреции ОП. Это в свою очередь приводит к увеличению секреции ОК, вазопрессина, ДА и ГнРГ, что проявляется в снижении уровня ПЛ, появлению молозива, повышении уровня ФСГ. Следующая за этим пролиферация гранулезы в фолликулах увеличивает концентрацию Э в крови, повышает секрецию ОП и активизирует импульсную секрецию ГнРГ.
При беременности хорион начинает секретировать во все увеличивающихся количествах хорионический гонадотропин (ХГ). Являясь аналогом ЛГ, он оказывает стимулирующее влияние на секрецию ОП, повышенный уровень которых надежно блокирует секрецию ГнРГ. ОП также блокируют дофаминовую систему, что проявляется в повышении секреции ПЛ, а так же блокируют выделение ОК.
Во внутриутробном периоде, как известно, после 11 — 12 недели возрастает секреция фетальных гонадотропинов. Это происходит в результате снижения секреции ХГ плацентой и снятия блока с гипоталамуса плода. Результатом повышенной секреции гонадотропинов является пролиферация гранулезы и формирование фолликулов в яичниках плода. Но увеличивающаяся продукция плацентой половых стероидов блокирует затем гипоталамус плода, секреция фетальных гонадотропинов снижается и к моменту рождения уровень их очень низок. В период новорожденности уровень половых стероидов снижается. Соответственно уровень гонадотропинов повышается.
В период полового созревания можно предположить следующие изменения. Известно, что в период детства секреции гонадотропинов и пролактина находятся на очень низком уровне. К началу полового созревания уровень их начинает повышаться. Импульсная же секреция гонадотропинов появляется вначале во время сна. Известно также, что во время сна повышается секреция пролактина. Учитывая эти факты, на основании вышеприведенной схемы можно предположить следующий ход событий. Одновременное угнетение секреции ГнРГ и ПЛ в период детства может происходить в результате повышенной секреции ДА. Повышение секреции ДА в период детства оправдано, т. к. ДА стимулирует продукцию гормона роста. Можно предположить, что эта секреция в процессе созревания нервной системы начинает снижаться, достигая к началу полового созревания уровня, когда блок с гипоталамуса снимается и начинается тоническая секреция ГнРГ, что проявляется повышением секреции ФСГ как легковысвобождающейся фракции гонадотропинов и повышением секреции ПЛ. Видимо, снижением уровня ДА можно объяснить остановку роста при преждевременном половом созревании. Постоянное воздействие ГнРГ на гипофиз стимулирует непрерывную секрецию гонадотропинов, препятствуя накоплению их в гонадотрофах. Можно предположить, что во время сна уровень ОП повышается (самостоятельно или через повышение ДА или серотонина), блокируя секрецию ГнРГ, что способствует накоплению гонадотропинов в гипофизе, в частности, ЛГ. Этому же способствует все повышающаяся секреция яичниками эстрогенов. Повышение запаса ЛГ в гонадогрофах и нарастание секреции эстрогенов в конце концов приводит к включению механизма импульсной секреции гонадотропинов, описанного выше. Этот процесс можно считать зеркальным отображением изменений, происходящих под действием синтетических аналогов ГнРГ, вводимых в непрерывном режиме.
Подобным образом можно объяснить и изменения при менопаузе: отсутствие гормонального стимула со стороны яичников и возрастные изменения нервной системы (снижение ДА) приводят к снижению секреции ОП и прекращению импульсной секреции ГнРГ, что проявляется во все увеличивающейся секреции ФСГ.
Известно, что обладающий выраженным рвотным эффектом апоморфин является опиатным и дофаминовым агонистом, а противорвотные препараты галоперидол и метоклопрамид — антагонисты дофаминовых рецепторов. При беременности дофаминовая система блокирована, что проявляется гиперпролактинемией. Известно также, что ДА стимулирует секрецию ОП. На основании вышеприведенных фактов можно сделать вывод, что причиной рвоты беременных является чрезмерная секреция ОП, вызванная влиянием ХГ.
Этим объясняется тошнота и рвота при приеме опиоидных аналгетиков, эстрогенов, дофаминомиметиков. При пузырном заносе уровень ХГ очень высок, что сопровождается неукротимой рвотой. При снижении уровня ХГ (угроза прерывания беременности) симптомы рвоты исчезают.
При раздражении соска (кормление), влагалища и шейки матки (коитус, роды) рефлекторно происходит повышение секреции окситоцина. Так как ОП тормозят секрецию ОК, в ответ на повышение его продукции секреция ОП повышается, что проявляется повышением уровня ПЛ в крови. Уровень ГнРГ, наоборот, снижается, чем объясняется контрацептивное действие грудного вскармливания.
Таким образом вышеприведенные факты можно считать подтверждением предположения о физиологическом значении звена гипоталамус-гипофиз как структуры, предназначенной для изгнания созревших продуктов жизнедеятельности репродуктивной системы - яйцеклетки и плодного яйца. Созревание обеспечивается сложной системой блокировки гипоталамических структур. Активизация же изгоняющей ситемы осуществляется изменением гормонального сигнала.
Подтверждением этого предположения могут служить и анатомические особенности этой структуры: единство паравентрикулярных и супраоптических ядер с задней долей гипофиза; различное эмбриональное происхождение передней и задней долей гипофиза; нахождение дугообразного ядра на пути окситоцинового тракта; наличие портальной системы кровообращения с возможностью обратного тока крови; отсутствие в срединном возвышении гемато-энцефалического барьера и наличие промежуточной доли гипофиза как своеобразного инактиватора опиоидов, поступающих сюда через портальную систему из гипоталамуса.
ЧАСТЬ ТРЕТЬЯ
Гормональные взаимоотношения между различными отделами репродуктивной системы женщины формируются в эмбриогенезе. Для анализа этих взаимоотношений как отношений между половыми клетками и организмом сведем данные о сроках закладок, стадиях развития и уровнях гормонов в период эмбриогенеза в одну таблицу (см. рис.2).
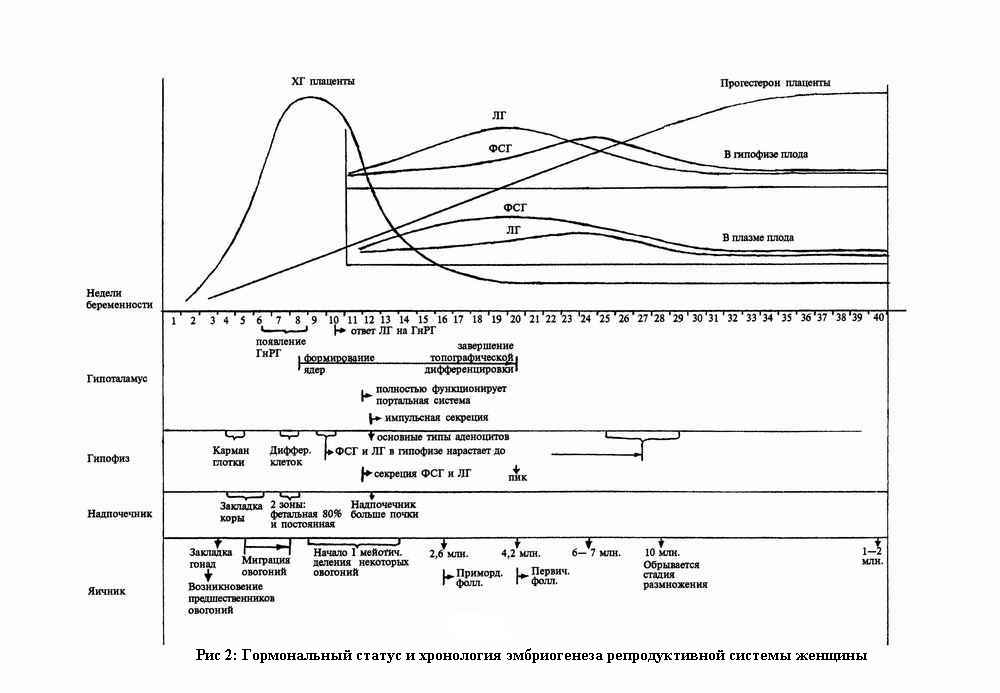
В эмбриогенезе половой системы обращает на себя внимание определенная связь закладок надпочечников и гонад. Известно, что, во-первых, они формируются в непосредственной близости друг с другом; во-вторых, биохимия синтеза гормонов очень схожа; в-третьих, закладка коры надпочечников происходит раньше, чем в закладке гонад появляются овогонии; в-четвертых, овогонии мигрируют из желточного мешка в область заладки гонад с помощью амебовидных движений (целенаправленность движения половых клеток позволяет предположить хемотаксис как стимулирующее влияние).
Учитывая вышеизложенное и принимая во внимание значение надпочечников в период внутриутробного развития, можно предположить, что надпочечники (в частности, закладка коры), являются источником хемотаксического стимула для овогоний.
Это объясняет обнаружение в области ворот яичников надпочечниковой ткани и обнаружение в ткани надпочечников так называемых «заблудившихся» гоноцитов.
Известно, что после периода миграции начинается период бурного размножения овогоний. К этому моменту формируется гипоталамо-гипофизарная система плода. После формирования портальной системы начинается импульсная секреция гонадотропных гормонов плода, уровень которых повышается после снижения продукции плацентой хориогонина. Можно предположить следующий ход событий. Размножение овогоний происходит без влияния гонадотропных гормонов плода, но формирование фолликулов и, как следствие, остановка мейотического деления происходит под влиянием гормонов гипофиза плода. Осуществляется изоляция овогоний. Но к 28 неделе беременности уровень гонадотропных гормонов плода снижается в связи с увеличением продукции плацентарных половых стероидов. Начинается массовая атрезия фолликулов. Этот процесс идет примерно с такой же скоростью, с какой шло увеличение числа фолликулов до 28 недели, и к моменту рождения остается 1-2 млн. фолликулов, а к периоду полового созревания — около 400 тысяч.
Что же происходит в зрелом яичнике ? Почему в каждом овариальном цикле процесс созревания начинается у определенного количества (пула) овоцитов? И почему в предовуляторный период вступает лишь один фолликул, а остальные подвергаются атрезии?
Известно, что ингибирование мейоза обусловлено составом внутренней среды фолликула и мейоз возобновляется в овоците: при аспирировании из зрелого фолликула и помещении в солевой раствор; при дегенерации гранулезных клеток; перед овуляцией.
Известно также, что ранним признаком атрезии являются возобновление мейоза и экструзия по меньшей мере первого полярного тельца. А гипофизэктомия у 100-дневных плодов макак резусов вызывала массовую гибель овоцитов и атрезию фолликулов в яичниках.
Следовательно, инициация мейоза у зрелого овоцита приводит к овуляции, а у незрелого - к атрезии. Можно предположить, что возобновление мейоза у зрелого овоцита инициирована самим овоцитом в процессе созревания, а возобновление мейоза у незрелого овоцита - это следствие инициации извне.
В таком случае в зрелом яичнике можно предположить следующее. После овуляции резкое снижение уровня гонадотропных гормонов провоцирует начало процесса созревания овоцитов. Этот процесс начинается не во всех фолликулах, а лишь в тех, в которых, по-видимому, овоциты вступили в мейоз раньше других (если предполагать, что овогонии мигрировали, размножались и вступали в I фазу мейоза не одновременно). Овоциты созревают до определенной стадии, когда в одном из них возобновляется мейоз. Экструзия первого полярного тельца вызывает изменение состава внутренней среды фолликула, которое влияет на внутреннюю среду остальных незрелых фолликулов этого пула и приводит к преждевременному возобновлению в них мейоза, гибели овоцитов и атрезии этих фолликулов.
Таким образом можно предположить, что надпочечники способствуют проникновению в эмбрион овогоний, а гипоталамо-гипофизарная система в течении последующей жизни изгоняет их из организма женщины. В этом, видимо, и заключается принцип функционирования репродуктивной системы: овогонии как клетки внеэмбрионального происхождения являются по отношению к плоду инородными клетками. Этим можно объяснить их массовую гибель — с 10 млн в 28 недель их остается к моменту рождения 1—2 миллиона, к моменту полового созревания - 400 тысяч, и в каждый овариальный цикл их гибнет сотни на один созревший овоцит.
Оценка функции репродуктивной системы как результата взаимоотношения между половыми клетками и организмом-носителем может помочь понять этиологию многих проблем акушерства и гинекологии.
ЛИТЕРАТУРА: