Дмитрий Марфунин
"Об аутоиммунном гепатите"
На главную
Аутоиммунный гепатит (AIH) чаще поражает женщин (70-80%) [12,25]. Различают несколько типов AIH, из них тип 2 отличается типично молодым возрастом начала [1]. Тип 2 AIH замечен преобладающе у детей и молодых женщин [16]. В Европе с типом 2 AIH до 80% пациентов – дети. Самые общие симптомы – усталость и миалгия, но 34% пациентов при начале являются бессимптомными. В дальнейшем 79% бессимптомных становятся симптомными [9]. Распространены артриты и артралгии [16]. При AIH наблюдается сочетание с различными аутоиммунными нарушениями, включая АИТ, язвенный колит, болезнь Грейвса, тип I диабет, ревматический артрит, глютеновую болезнь [9,15,16]. Иммунные болезни могут развиться в любое время в течение AIH [9]. Но частота аутоиммунных нарушений, которые специфически поражают печень, таких, как AIH, относительно низка по сравнению с другими аутоиммунными болезнями [13]. У женщин AIH может проявиться во время беременности и в ранний послеродовой период [16].
Гистологически AIH характеризуется плотным портальным инфильтратом моноцитарных клеток, вторгающимся в ограничивающую пластину, то есть резко разделяющую гепатоциты границу, который окружает портальную триаду и проникает в окружающую паренхиму. Этот инфильтрат включает Т, В-лимфоциты, макрофаги и плазматические клетки [15,26]. У пациентов со спонтанной или фармакологически вызванной ремиссией гистологические изменения могут вернуться к норме, фиброз может регрессировать или исчезнуть [16].
При AIH типа 2 определяются анти почек печени микросом антитела (LKM-1). Эти антитела распознают короткой длины эпитоп в пределах фермента цитохрома монооксидазы P4502D6 (CYP2D6), который составляет главный человеческий антиген при типе 2 AIH [1,4,9,13]. У LKM-1 позитивных пациентов AIH имеет тенденцию возникать более остро и в младшем возрасте [25]. Главный эпитоп CYP2D6-193-212 также признается у 50% пациентов с LKM-1 положительной инфекцией вирусным гепатитом С (HCV) [26]. Но хотя титры анти-CYP2D6 антител у пациентов с инфекцией HCV и типа 2 AIH подобны, все же картирование эпитопов показало, что имеются существенные различия относительно эпитопов, распознаваемых LKM-1s антителами HCV-инфицированных пациентов и LKM-1s антителами пациентов AIH-2, что указывает на расходящиеся пути, приводящие к расстройству толерантности к CYP2D6 в этих двух ситуациях [12,13].
У 50% пациентов с LKM-1 положительными сыворотками обнаруживают анти-LC-1 антитело, которое реагирует со специфическим для печени 58-kD метаболическим ферментом formiminotransferase cyclodeaminase в титрах, которые коррелируют с активностью болезни типа 2 AIH. Это антитело было также описано у 10% пациентов с типом 2 AIH, где оно представляло единственный обнаруживаемый маркер болезни [12,15].
Генерация анти-CYP2D6 антител по существу не достаточна, чтобы вызвать аутоиммунное повреждение печени [13]. Может быть серонегативный AIH, подчеркивая возможность отсутствия циркулирующих аутоантител при AIH [16]. Помимо подавляющего В клетка/антитело-опосредованного иммунного ответа на антигены печени, агрессивный CD8 Т клеточно-опосредованный процесс мог быть вовлечен в патогенез AIH-2 [13]. CD4 Т клетки от пациентов с типом 2 AIH распознают семь областей CYP2D6, пять из которых также распознаются CD8 Т клетками. Циркулирующие Т клетки, специфические для аутоантигенов печени, найдены и у нормальных субъектов, но при AIH их частота 10-кратно увеличивается. При AIH-2 наблюдается низкий уровень пролиферации Т клеток, экспрессирующих маркер CD8. Среди Т клеток существуют CD4+ Т клетки, конститутивно экспрессирующие рецептор IL-2 (CD25, Т-regs), которые контролируют иммунные ответы, предотвращая быструю пролиферацию и функции эффекторно аутореактивных Т клеток. У здоровых эти клетки представляют 5-10% общего числа периферийных CD4+ Т клеток, при AIH их число сокращается, что, как предполагают, способствует проявлению AIH [26].
Показано, что фермент formiminotransferase cyclodeaminase (FTCD), кроме своего участия в деградации гистидина, вовлечен во взаимодействие аппарата Гольджи с микроканальцевым цитоскелетом. FTCD стимулирует взаимодействие между элементами Гольджи и vimentin промежуточных филаментов (IF) и действует как молекулярный мост, связывающий мембраны Гольджи и vimentin [10]. Как известно, микротрубочки цитоскелета участвуют во внутриклеточном транспорте. Учитывая анти-CYP2D6 антитела, анти-LC-1 (FTCD) антитела, а также присутствующие иногда анти-актиновые антитела, складывается впечатление, что иммунная ткань стремится подавить как ферментативную активность CYP2D6, так и внутриклеточный транспорт в гепатоцитах. Вышеперечисленные факты позволяют думать о том, что CYP2D6 может играть существенную роль в функционировании лимфоидной ткани.
В лимфоцитах периферической крови ген CYP2D6 типично экспрессировался на самых высоких уровнях [17,23]. Хотя CYP2D6 mRNA содержание было измерено во всех образцах лимфоцитов периферической крови [11,23], оно соответствует примерно одной десятой экспрессируемого в человеческой печени [17]. Содержание CYP2D6 в человеческой печени оценивается приблизительно 7 pmol/mg общего белка печени. При условии, что экспрессия белка лимфоцита является 2 порядками ниже, приблизительно 70 fmol CYP2D6 в мг белка могло бы присутствовать в лимфоцитах. Но белок CYP2D6 не был обнаружен в лимфоцитах. Отказ обнаружить активность CYP2D6 в человеческих инкубированных лимфоцитах означает, что или функциональный CYP2D6 полностью отсутствует, или количество CYP2D6, экспрессируемого в лимфоцитах, должно быть более чем 100-кратно ниже, чем в печени [19], что находит подтверждение в недавнем исследовании, показавшем, что по сравнению с печенью концентрация Р450 в периферической крови мононуклеарных клетках в 20-2000 раз ниже [23]. О ферменте можно судить косвенно, так кодеин, добавленный к мононуклеарным клеткам крови человека, был деметилирован в морфин, указывая на присутствие функционально активного CYP2D6 [2]. Учитывая агрессию иммунной ткани, направленную на фермент CYP2D6, и согласно принципу обратной связи, можно предположить, что или продуктом катализируемой CYP2D6 реакции являются вещества, негативно влияющие на иммунную ткань, или же в катализируемой CYP2D6 реакции расходуется субстрат, необходимый для жизнедеятельности иммунной ткани.
CYP2D6 участвует в метаболизме многих экзогенных и эндогенных веществ. Среди эндогенных биологически активных веществ обращает на себя внимание группа индолэтиламинов. Так известно, что CYP2D6 практически не метаболизирует эндогенные серотонин, мелатонин, 6-метокситриптамин [30]. Но эндогенные 5-метоксииндолэтиламины, полученные из серотонина [5-гидрокситриптамина (5-НТ)], были идентифицированы как высокоаффинные субстраты CYP2D6, который катализирует их О-деметилирование с высокой ферментативной способностью и спецификой [31].
Чтобы лучше представить специфику взаимоотношений лимфоцитов и серотонина, необходимо сделать небольшое отступление.
Лимфоциты и серотонин
Известно, что основная часть 5-НТ в организме продуцируется и секретируется энтерохромаффинными клетками (ЕС) желудочной и кишечной слизистой оболочки [5]. Предполагается взаимодействие между 5-НТ секрецией ЕС и лимфоцитами в слизистой оболочке кишки [27]. 5-НТ быстро захватывается тромбоцитами и в меньшей степени лимфоцитами [5]. 5-НТ концентрация может быть регулируема его транспортом в моноциты, макрофаги, дендритные клетки и лимфоциты через активную систему 5-НТ захвата [5-НТ транспортер (5-НТТ)] [6].
В лимфоцитах обнаружено mRNA кодирование 5-НТ-1В, 5-НТ-1F, 5-НТ-2А, 5-НТ-2В, 5-НТ-3, 5-НТ-6 и 5-НТ-7 субтипов рецепторов. mRNA для 5-НТ субтипов рецепторов были обнаружены и в покоящихся человеческих лимфоцитах [5]. Наивные Т клетки преобладающе экспрессируют тип 7 5-НТ рецептора и экспрессия этого белка существенно увеличена при активации Т клетки. 5-НТ передача сигнала через 5-НТ 7 рецептор может способствовать ранней активации Т клетки [18].
5-НТ стимулирует продукцию INF-гамма NK клетками человека. 5-НТ индуцирует продукцию IL-16, хемотаксического фактора в CD8+ Т клетках [5]. 5-НТ стимулирует транскрипцию NF-кВ в спленоцитах, что связано со стимуляцией 5-НТ пролиферации лимфоцитов [6]. 5-НТ индуцирует быстрое фосфорилирование внеклеточными сигналами регулируемой киназы-1 и 2 (ERK1/2) и IкВ-альфа в наивных Т клетках [18]. 5-НТ может дифференцированно модулировать LPS -индуцированную продукцию цитокинов в человеческих лимфоцитах на посттранскрипционном уровне, а также ингибировать продукцию этими клетками TNF-альфа [5]. 5-НТ увеличивал ex vivo экспансию линии крови CD34+ гематопоэтических стволовых клеток и клеток предшественников [28]. Таким образом можно думать о том, что 5-НТ играет существенную роль в активации Т клеток. Было предположено, что 5-НТ выступает в качестве медиатора двунаправленных взаимодействий между нервной и иммунной системами [28].
Резидентные в периферийных тканях дендритные клетки (DCs) обладают механизмами для специфического захвата и регулируемого выпуска 5-НТ, могут захватывать 5-НТ, выпущенный активированными тромбоцитами, дегранулированными тучными клетками или симпатическими нервами, изолировать его во внутриклеточных пузырьках и выпускать сохраненный 5-НТ путем Са2+-вызванного экзоцитоза. Предполагается также, что DCs могут захватывать 5-НТ из окружающей микросреды или непосредственно от активированных Т клеток и передавать его наивным Т клеткам [21].
Активация Т клеток требует зоны адгезии, прямого контакта с антигенпредставляющими клетками, обычно известного как иммунный синапс. Иммунный синапс максимизирует физическое взаимодействие между DCs и лимфоцитами и формирует устойчивую платформу для вербовки и концентрации сигнальных молекул. Иммунный синапс может также поддерживать регулируемую секрецию лабильных молекул, которые могли способствовать передаче сигналов между DCs и лимфоцитами. Фокальный выпуск 5-НТ DCs в синаптических соединениях с Т клетками может подавить продукцию сАМР и таким образом облегчить активацию и дифференцирование наивных Т клеток. Хотя лабильные и физиологические уровни 5-НТ в сыворотке редко превышают наномолярные концентрации, выпуск 5-НТ в синаптическое пространство между DCs и Т клетками произвел бы ограниченную высокую концентрацию 5-НТ, способную действовать на короткие расстояния и сигнальные 5-НТ рецепторы [21].
Идентифицирована экспрессия типа 1 триптофангидроксилазы (ТРН-1) активированными Т клетками, что предполагает способность к продукции собственного 5-НТ [18,21]. IFN-гамма вызывает выпуск 5-НТ из массы лимфоцитов периферийной крови [18]. Истощение внутриклеточного запаса 5-НТ уменьшило способность селезеночных Т клеток пролиферировать и экспрессировать IL-2 R в ответ на конкавалин А (ConA). Ответ восстанавливался добавлением 5-НТ. Пролиферация Т клеток на ConA и IL-2 уменьшалась присутствием антагонистов к 5-НТ-2 рецептору. Истощение клеточных запасов 5-НТ привело к ослаблению бластогенеза Т клеток, восстановленному дополнением 5-НТ [29].
Показано, что покоящиеся и стимулируемые лимфоциты человека синтезируют и выпускают значительное количество мелатонина. Описаны высокоаффинные мелатонин-связывающие участки в лимфоцитах человека [3]. Рецепторы мелатонина являются присущей особенностью популяций активированных лимфоцитов человека [14].
Мелатонин активирует Т хелперы клетки, увеличивая продукцию IL-2, а также и IL-12, ведя дифференцирование Т клетки к фенотипу Th1 и вызывая увеличение продукции IFN-гамма [3]. Мелатонин и 5-метокситриптамин (5-МТА) могут влиять на пролиферативный ответ лимфоцитов. Так, мелатонин ингибировал, а 5-МТА увеличивал ConA-индуцированное вхождение тимидина в лимфоциты. Ко-инкубация лимфоцитов с ConA и мелатонином привела к ингибиции специфичного связывания иода, но с 5-МТА не отличалась от контроля [14]. Таким образом 5-МТА проявляет свойства агониста 5-НТ. Есть противоречивые данные о синтезе мелатонина в стимулируемых и нестимулируемых лимфоцитах, так как при стимуляции лимфоцита наблюдается увеличение ферментативной активности серотонин-N-ацетилтрансферазы (NAT), конвертирующей 5-НТ в N-ацетилсеротонин, но активность HIOMT (гидроксииндол-О-метилтрансферазы) была значительно ингибирована [3].
Известно, что 5-НТ при вхождении в клетку метаболизируется тремя путями. Путем окислительного дезаминирования с помощью фермента моноаминоксидазы (МАО) в 5-гидроксииндол-3-ацетат и далее в виде неактивных продуктов удаляется из клетки. Путем N-ацетилирования с помощью NAT в N-ацетилсеротонин. И путем метилирования в 5-МТА. Далее N-ацетилсеротонин путем метилирования с помощью HIOMT конвертируется в мелатонин, а 5-МТА путем окислительного дезаминирования с помощью МАО конвертируется в 5-метоксииндол-3-ацетат и далее в виде неактивных продуктов удаляется из клетки (см. схему).
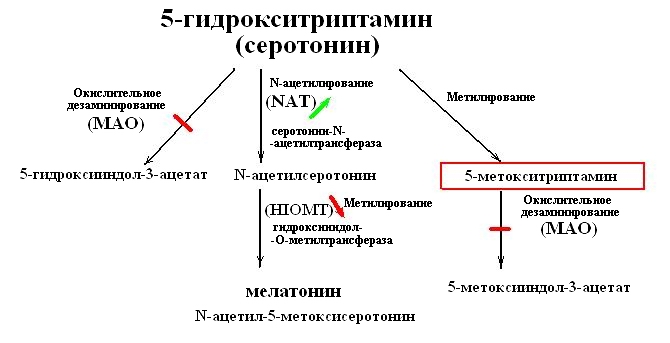
Как известно, существует 2 основных типа моноаминооксидазы – МАО-А и МАО-В. МАО-А предпочтительно окисляет 5-НТ и норадреналин, тогда как МАО-В – РЕА (фенилэтиламины – катехоламины) [22,24]. МАО-А-КО мыши (нокаутные по МАО-А) увеличили уровни 5-НТ, тогда как МАО-В-КО мыши (нокаутные по МАО-В) увеличили уровни РЕА. То есть у МАО-А и В есть различные функции в метаболизме медиаторов [22]. И МАО-А и МАО-В вовлечены в завершение действия аминов медиаторов [24].
Показано, что тромбоциты, лимфоциты и некоторые лейкоциты экспрессируют МАО-В, тогда как никто из них не экспрессирует обнаруживаемую МАО-А. То есть лимфоциты экспрессируют только МАО-В [24].
Из всего следует, что 5-НТ, попадая в лимфоцит, не может окисляться в 5-гидроксииндол-3-ацетат, а его производное 5-МТА не может окисляться в 5-метоксииндол-3-ацетат. Но известно, что 5-МТА является эндогенным субстратом CYP2D6, который является относительно высоко специфичной, высокоаффинной, высокопроизводительной 5-метокситриптамина О-деметилазой [30]. Можно думать о том, что именно CYP2D6 в активированном лимфоците завершает действие медиатора 5-НТ.
Продолжение
Синтез и деградация 5-НТ – активный процесс, с общим пулом 5-НТ, заменяемым каждые 24 часа [28]. Секреция 5-НТ ЕС стимулируется посредством передачи сигналов от бета-адренергических и РАСАР рецепторов, а ингибируется посредством нервных медиаторов САВА и ацетилхолина [20]. С другой стороны, 5-НТ, выпущенный кишечными ЕС, активирует 5-НТ-3 рецепторы на вагусных центростремительных волокнах, в конечном счете стимулируя выброс ацетилхолина [32]. Все это выглядит вполне логично, так как формируется классическая петля отрицательной обратной связи, регулирующая секрецию 5-НТ. При снижении уровня ацетилхолина его ингибирующее влияние уменьшается и может быть причиной увеличения уровня 5-НТ. В норме энтероциты, которые экспрессируют транспортер серотонина (SERT,5-НТТ), в значительной степени ответственны за завершение действия 5-НТ, удаляя его из интерстициального пространства [8]. При увеличении уровня 5-НТ можно предположить, что к процессу инактивации 5-НТ может активно подключиться печень, причем энзимотически метаболизируя его и его производные с помощью как МАО, так и CYP2D6.
Можно думать о том, что, попадая в наивную Т клетку, 5-НТ быстро метаболизируется (см. схему). Образующийся при его метилировании 5-МТА, очевидно, и является тем агентом, который активирует Т клетку, стимулируя ее пролиферацию и дифференцирование и обеспечивая ее бластогенез. Этим, видимо, можно объяснить положительный эффект от применения препарата 5-МТА (мексамин) как радиопротектора. Одновременно стимулируется продукция белка CYP2D6, необходимого для инактивации 5-МТА.
Чрезмерная инактивация 5-МТА посредством CYP2D6 печени, возможно, и вызывает агрессивный ответ лимфоидной ткани, направленный в первую очередь на подавление активности CYP2D6 печени.
Предполагается, что гонадотропины стимулируют активность ацетилхолинэстеразы в крови (см. «О миастении»), что может привести к снижению уровня ацетилхолина и, как следствие, повышению уровня 5-НТ. Возможно, что именно это обстоятельство могло быть причиной преобладания молодых женщин среди пациентов AIH-2. Так же, как и проявления AIH во время беременности и в раннем послеродовом периоде.
Предполагается, что у детей повышенный уровень продукции 5-НТ, что так же можно было бы считать причиной их более частого поражения AIH, чем взрослых.
Показано, что глютеновая болезнь довольно часто сочетается с AIH, особенно отмечается ее бессимптомность и возможность способствовать дисфункции печени [9]. Отмечено, что при глютеновой болезни увеличено число 5-НТ-содержащих ЕС, она была связана с увеличенным содержанием 5-НТ слизистой оболочки и значительно более высоким уровнем плазменного 5-НТ [7].
Итак, на основании вышеизложенного складывается впечатление, что поражение печени аутоиммунным гепатитом типа 2 можно было бы считать реакцией лимфоидной ткани на чрезмерную активность фермента CYP2D6 печени в результате повышения уровня серотонина в крови.
То, что при AIH-2 отмечается возможность спонтанной ремиссии, а также обнаружение у нормальных субъектов циркулирующих Т клеток, специфических для аутоантигенов печени, позволяет рассматривать AIH-2 как дизрегуляционный процесс.
ЛИТЕРАТУРА :
1. Berg PA, Klein R. Praxis ( Bern 1994). 2002 Aug 21; 91(34):1339-46
2. Boettcher C, Fischer W, Zenk MN . The Journal of Immunology. 2006, 176: 5703-5704
3. Carrilla-Veco A, Calvo JR, Abreu P, Lardone PJ, Garcia-Maurino S, Reiter RJ, Guerrero JM. The FASEB Journal express article 10.1096/fj.03-0694fje.Published online January 8, 2004
4. Christen U, Holdener M, Hintermann E. Autoimmun Rev. 2007 Apr;6(5):306-11
5. Cloez-Tayarani I, Petit-Bertron A-F, Venters HD, Cavaillon J-M. International Immunology, Vol.15, No.2, pp 233-240 February 2003
6. Cloez-Tayarani I, Ghangeux J-P. Journal of Leukocyte Biology. 2007;81:599-606
7. Coleman NS, Foley S, Dunlop SP, Wheatcroft J, Blackshaw E, Perkins AC, Singh G, Marsden CA, Holmes GK, Spiller RC. Clin Gastroenterol Hepatol. 2006 Jul; 4(7):874-88
8. Crowell MD. Br J Pharmacol. 2004 April;141(8)1285-1293
9. Czaja AJ. MedGenMed. 2006; 8(2):55
10. Gao Y-S, Sztul E. The Journal of Cell Biology. Volum 152, Number 5, March 5, 2001 877-894
11. Haas CE, Brazeau D, Cloen D, Booker BM, Frerichs V, Zaranek C, Frye RF, Kufel T. Eur J Clin Pharmacol. 2005 Sep; 61(8):583-93
12. Heneghan MA, McFarlane IG. Hepatology 2005 Volume 42, Issue 1, Pages 17-20
13. Holdener M, Hintermann E, Bayer M, Rhode A, Rodrigo E, Hintereder G, Iohnson EF, Gonzalez FJ, Pfeilschifter J, Manns MP, Herrath M. von G, Christen U. The Journal of Experimental Medicine, April 2008 Vol. 205, No.6, 1409-1422
14. Kanakchieva R, Kyurkchiev S, Kehayov I, Taushanova P, Kanchev L. J Neuroimmunol. 1995 Dec 31; 63(2):125-32
15. Krawitt EL. The New England Journal of Medicine January 5, 2006 Number 1 Volume 354:54-66
16. Krawitt EL. World J Gastroenterol 2008 June 7; 14(21):3301-3305
17. Krovat BC, Tracy JH, Omiecinski CJ. Toxicological Sciences 55, 352-360 (2000)
18. Leon-Ponte M, Ahern GP, O`Connell PJ. Blood, 15 April 2007, Vol. 109,No.8,pp.3139-3146
19. McConnachie LA, Phillips B, Bajpai M, Sheu DD, Ho RJY. DMD 31:1103-1107, 2003
20. Modlin IM, Kidd M, Pfragner R, Eick GN, Champaneria MC. The Journal of Clinical Endocrinology and Metabolism Vol. 91, No. 6 2340-2348 2006
21. O`Connell PJ, Wang X, Leon-Ponte M, Griffiths C, Pingle SC , Ahern GP. Blood, 1 February 2006, Vol. 107, No. 3, pp 1010-1017
22. Shih JC, Cheu K. Neurobiology (Br). 1999; 7(2):235-46
23. Siest G, Jeannesson E, Marteau J-B, Samara A, Marie B, Pfister M, Visvikis-Siest S. Drug Metabollism and Disposition, 2008 36:182-189
24. Thorpe LW, Westlung KN, Kochersperger LM, Abell CW, Denney RM. The Journal of Clinical Endocrinology and Metabolism Vol. 91, No. 6 2340-2348 2006
25. Vergani D, Mieli-Vergani G. Minerva Gastroenterol Dietol. 2004 Jun; 50(2): 113-23
26. Vergani D, Mieli-Vergani G. World J Gastroenterol. 2008 June 7; 14(21) :3306-3312
27. Yang GB, Lackner AA. J Neuroimmunol. 2004 Jan; 136(1-2): 46-9
28. Yanga M, Lia K, Nga PC, Chuena CKY, Laub TK, Chenge YS, Luic YS, Lia CK, Yuena PMP, Jamesd AE, Leea SM, Foka TF. Stem Cells Vol. 25 No. 7 July 2007. pp. 1800-1806
29. Young MRI, Kut JL, Coogan MP, Wright MA, Young ME, Matthews J. Immunology 1993 80 395-400
30. Yu AM, Idle JR, Herraiz T, Kupfer A, Gonzalez FJ. Pharmacogenetics 2003 Jun; 13(6):307-19
31. Yu AM, Idle JR, Gonzalez FJ. Drug Metab Rev. 2004 May; 36(2):243-77
32. Zhu JX, Wu XY, Owyang C, Li Y. J Physiol. 2001 February 1; 530 (Pt 3):431-442
В начало
На главную
© Дмитрий Марфунин